

|
||||||||||
Lintwurms het ’n indirekte lewensiklus. Dit beteken daar is twee gashere: ’n primêre gasheer, waarin die volwasse parasiet voorkom, en ’n tussengasheer, waarin ’n ander stadium, bekend as die metasestood, voorkom. Die lintwurm Taenia solium, met sy feitlik wêreldwye verspreiding, is inderdaad seker die bekendste parasiet wat mense besmet. Reeds so vroeg as 1550 v.C. is verwysings gevind na lintwurmbesmetting in mense. Daar is egter ’n groot aantal ander spesies wat tot dieselfde genus behoort. Spesies wat in Suid-Afrika in honde en katte voorkom, sluit die volgende in: T. hydatigena, T. multiceps, T. ovis ovis, T. pisiformis, T. serialis serialis, T. serialis brauni en T. taeniaeformis. Laasgenoemde kom in katte (Felidae) voor en die res in hondagtiges (Canidae). Die vraag ontstaan of dieselfde Taenia spp.-wurms wat in honde voorkom ook in “wilde” hondagtiges (soos jakkalse) en katagtiges (soos rooikatte) voorkom in Suid-Afrika en indien wel, tot watter mate.
Inligting (afmetings van aanhegtingstrukture) oor lintwurms uit honde is verkry uit ClinVet International (Edms.) Bpk. se argiefmateriaal. Lintwurmmateriaal is ook versamel uit jakkalse en rooikatte (Bloemfontein en omgewing in die Vrystaat) wat spesifiek vir die studie verwerk is.
Resultate toon dat daar wel oorvleueling is tussen die Taenia spp.-samestelling van honde en jakkalse (byvoorbeeld T. hydatigena). Die samestelling van spesies kan beïnvloed word deur faktore soos geografiese verspreiding en dieet. Verdere studies kan moontlik meer area-spesifieke opnames insluit vir beide gasheerspesies, met monsterneming uit meer geografiese areas (m.a.w. wat meer verteenwoordigend is van Suid-Afrika as geheel). Dit kan ook gekombineer word met ’n tipe sosio-ekonomiese studie om te kyk na die omvang en rol van informele slagpraktyke in veral landelike gebiede in die lewensiklus van Taenia spp.-parasiete.
’n Poging sal daarmee saam aangewend moet word om die graad van predasie deur jakkalse te kwantifiseer, die rol van honde in die lewensiklus te evalueer en dit te vergelyk met die voorkoms van die Taenia spp. in beide gashere. Verder wil dit voorkom of rooikatte lintwurms kan huisves wat eie is aan honde (T. hydatigena), asook dié wat eie is aan katte (T. taeniaeformis). Die verskynsel moet in verdere studies bevestig en verder bestudeer word.
Trefwoorde: Taenia sp.; lintwurm; hond; Canis familiaris;jakkals; Canis mesomelas; rooikat; Caracal caracal.
Parasitologists are most likely the only people that get excited by discussions on tapeworm! For most people the mere notion of tapeworm infections is gross, the first thought usually involving undercooked pork. Taenia solium may indeed well be the best-known tapeworm, yet this genus also comprises several other species. They all share similarities when it comes to the life cycle, which is described as being indirect. This implies that two hosts are involved: the first a final host containing the “adult” parasite (i.e. scolex behind which segments form and develop) and the second an intermediate host containing the larval stage. Once again T. solium serves as a good example. The “adult” worm may be 2 to 7 metres long and resides in the small intestine of infected humans. As proglottids develop from behind the scolex, the oldest, most developed proglottids are right at the end of the worm. These contain eggs and when fully developed the proglottid is expelled with the host faeces. Pigs ingest the proglottids and/or viable eggs which “hatch” to form oncospheres. The latter penetrates the intestinal wall and is distributed to muscle tissue (i.e. “measles”) through blood and lymph flow. Should a human ingest viable eggs, oncosphere development would progress as it would in an intermediate host, which may lead to cyst formation in various tissues (e.g. liver and brain). As mentioned before, several species of Taenia exist, but for the purposes of this manuscript the focus will be on those found in cats and dogs. Work by Verster (1969) and a subsequent review by Loos-Frank (2000) implicates the following species within a South African context: T. hydatigena, T. multiceps, T. ovis ovis, T. pisiformis, T. serialis serialis, T. serialis brauni and T. taeniaeformis. The latter occur in felids only, while the rest are found in canids. All the representatives from this genus possess two sets of hooks used for attachment. Measurements of these hooks are also used for taxonomic purposes (i.e. species identification).
The question arises whether the same Taenia spp. that occurs in dogs also occurs in wild canids (e.g. jackal) or felids (e.g. caracal) and if so, to what extent. Archaeological evidence indicates that domestic dogs (Canis familiaris) are not endemic to Africa. They were brought into Egypt from Europe approximately 7 000 years ago (Gallant 2004) and distributed through Africa from there. The jackal (Canis mesomelas) often shares the same parasite and diseases with domestic dogs (Price en Karstad 1980; Walton en Joly 2003; Loveridge en Nel 2004).
Jackals are omnivorous, opportunistic feeders and will both hunt small prey and feed on available organic material (Skinner en Smithers 1990; Walton and Joly 2003). Caracals (Caracal caracal) are predominantly hunters and also feed on small to medium-sized prey (Skinner en Smithers 1990).
During this preliminary survey, ClinVet International (Pty) Ltd archive material was examined to investigate what Taenia spp. is commonly found in domestic dogs in South Africa (mainly from the Grahamstown area in theEastern Cape). With the aid of the Free State Sandveld branch of the South African Hunters and Game Conservation Association (SAHGCA) a number of jackal and caracal carcasses were obtained from the Bloemfontein area (Free State) and examined for the presence of tapeworm.
Obviously the geographical areas are very limited and not at all representative of conditions in greater South Africa. Furthermore, the amount and quality of material and hence data was not suitable for intensive statistical analyses. It was, however, adequate for defining and investigating the following aims in this pilot study: (1) Identify what Taenia spp. occur in dogs in South Africa and compare that to species found in jackal. (2) Identify Taenia spp. found in caracal and compare that to species found in dogs and jackal. (3) Discuss variation in measurements of hooks and discuss implications for identification of specimens examined.
Measurements of hooks from Taenia spp. recovered from dogs were obtained from the archives of ClinVet International (Pty) Ltd. Taenia spp. tapeworm were also collected directly from the small intestines of jackal and caracal. Visible worms were removed and the mucosa was lightly scraped and strained, and the washing examined for scoleces. All parasites were preserved in 10% formalin. Subsequently the rostellum containing hooks were carefully cut off from each scolex and mounted in lactophenol (alternatives for use in future studies are Berlese’s or Hoyer’s media). With the use of two needles the rostellum was manipulated to obtain a suitable orientation before placing a cover slip over it. The edges of the cover slip were sealed with clear Cutex. The respective lengths of the long and short hooks (five of each for each parasite examined) were measured using a measuring eye piece. The average lengths were subsequently used for identification purposes.
Comparatively few of the specimens collected from 18 jackals (14 out of 112 Taenia spp. worms collected) were suitable for purposes of species identification. This was attributed to the fact that degradation of soft tissues in which the hooks are embedded takes place very rapidly, resulting in hooks being lost as carcasses could not be collected immediately. In future studies, intestines should be collected immediately in the field, placed on ice and transported to the laboratory. Out of three Taenia spp. specimens, collected from three caracal, only two were in a suitable condition. Measurements for 24 Taenia spp. specimens from eight dogs were obtained from the ClinVet archives. Subsequent examination, comparison and discussion were restricted to Taenia spp. previously recorded from South Africa as recorded by Verster (1969) and later reviewed by Loos-Frank (2000). The measurement ranges recorded by Verster (1969) were graphically presented for purposes of comparison, but additional measurements and ranges were also tabulated. Results indicate that the majority of Taenia spp. previously collected from dogs belonged to the species T. hydatigena. This is not surprising, as T. hydatigena is a common, cosmopolitan species that has been found in several intermediate and final host species.A number of recovered Taenia spp. specimens are likely to be T. ovis ovis. Both these Taenia spp. also use sheep as an intermediate host. It is very likely that infected sheep tissues (e.g. muscle for T. ovis and liver for T. hydatigena) were removed during rural slaughter practices and then thrown to the dogs to be eaten (or discarded and then later found by dogs).There was also one specimen from the T. multiceps / T. serialis serialis “measurement range complex”and two others that fell outside the measurement ranges defined by Verster (1969). This largely corresponded to what was found in jackal. However, only two specimens are suspected to be T. hydatigena,while the majority fell into the T. multiceps / T. serialis serialis / T. ovis ovis “measurement range complex”. The hook measurements of three specimens corresponded to those previously recorded for T. serialis brauni. Infection statistics calculated for Taenia spp. from jackal were as follows: Prevalence = 22,22%; Mean intensity = 28,00; Mean abundance = 6,22. While not corresponding to measurement value ranges provided by Verster (1969), it seems likely that at least one Taenia spp. collected from caracal belongs to the species T. taeniaeformis, a parasite of felids using rodents as intermediate hosts. The other Taenia spp. specimen collected appears to be T. hydatigena.
From this preliminary study caracal may thus appear to be infected with Taenia spp. tapeworms associated with both canids and felids. This should, however, be further investigated and confirmed in future studies. Infection statistics calculated for Taenia spp. from caracal were as follows: Prevalence = 66,67%; Mean intensity = 1,50; Mean abundance = 1,00. Variation in measurements of structures for purposes of species identification can be influenced by a number of factors, which may include researcher experience and measurement methodology. In other groups, such as monogenean worms, and more specifically Gyrodactylus spp., morphometric variation became such a confounding factor that the majority of new descriptions are accompanied by molecular analyses. Such protocols have been developed and are well recorded for Taenia spp.
Future studies should employ an integrated approach that would include experimental infections with materials of known species origin, traditional morphometric approaches and molecular analyses.
A number of deductions were made following completion of this pilot study. When examining Taenia spp., material should be as fresh as possible to avoid losing the attachment hooks. The use of different clearing agents (e.g. Hoyer’s medium versus lactophenol) should be investigated in future studies.
There was an overlap in the Taenia spp. fauna of jackal and dogs. As species composition is affected by factors such as geographical distribution and diet, future studies should focus on larger and more representative sampling areas. This should be coupled with socio-economic studies to examine the types of rural slaughtering practices in use and how these further the parasite life cycle. Furthermore, the extent of predation on sheep by jackal should be quantified, after which parasite burdens between jackal and domestic dogs from the same area can then be compared.
It would also appear as if caracal can harbour Taenia spp. found in both canids and felids. This should be further investigated in future studies. Further morphometric studies are also required where various measurement methodologies are compared, combined with experimental and molecular approaches, as previously stated.
Keywords: Taenia sp.; tapeworm; dog; Canis familiaris; jackal; Canis mesomelas; caracal; Caracal caracal.
“Wurmstukkies. Die plat wit tipe, wat ook as proglittiede bekend staan. Stoelgange kan deurspikkel wees (Figuur 1) en selfs langsaam alleen uit ’n rektum kan kruip ...” Die beskrywing klink nie net soos iets uit ’n grufilm nie, maar lok ook dieselfde grilreaksie in die meeste mense uit wat met die idee van lintwurms konfronteer word. Helaas, dit is inderdaad nie die tipe onderwerp wat enige “normale” persoon vrywillig en met entoesiasme sal bespreek nie. Gelukkig is parasitoloë nie “normale” mense nie.

Figuur 1. Hondefeses met lintwurmproglottiede duidelik sigbaar. Die kleiner proglottiede wat soos ryskorrels lyk, is dié van Dipylidium caninum. Die groter, platter proglottiede is dié van Taenia spp., vermoedelik Taenia hydatigena.
(Foto: Petro Swart, ClinVet International)
Wanneer mense die woord “lintwurm” hoor, het hulle eerste gedagte gewoonlik iets te make met rou varkvleis. Die lintwurm Taenia solium, met sy feitlik wêreldwye verspreiding, is inderdaad seker die bekendste parasiet wat mense besmet.
Reeds so vroeg as 1550 v.C. is verwysings gevind na lintwurmbesmetting in mense (Grove 1990). In teenstelling met besmetting met volwasse wurms, wat maklik behandelbaar is, is die vorming van siste in menslike weefsel (soos die brein, lewer en longe) meer skrikwekkend en baie gevaarliker. Nog ’n lintwurm waarvan die volwasse stadium net mense besmet, is T. saginata. Soos T. solium se besmetlike tussenstadium (m.a.w. besmetlik vir die mens as primêre gasheer) in die vark (tussengasheer) voorkom, so kom T. saginata se besmetlike stadium in beesvleis voor (Minozzo, Gusso, de Castro, Lago en Soccol 2002). Daar is ook nog ’n derde vorm geïdentifiseer (T. asiatica) wat mense besmet en eienskappe van beide T. solium en T. saginata toon (Eom en Rim 1992; Eom, Rim en Geerts 1992; Galán-Puchades en Fuentes 2000; Wang, Chung, Lu en Fan 2000). Verwys na Flisser, Viniegra, Aguilar-Vega, Garza-Rodriguez, Maravilla en Avila (2004) vir ’n oorsig van die drie spesies.
Alhoewel die lintwurmspesies uit die aard van die primêre gasheer bekendheid verwerf het onder die algemene publiek, besef min mense dat daar ’n groot aantal spesies in dieselfde genus is wat ’n groot verskeidenheid werweldiere besmet. Hoberg, Alkire, De Queiroz en Jones (2001) spekuleer dat Taenia spp.-lintwurms wat mense besmet, ontstaan het in Afrika nadat moontlike gasheerwisseling plaasgevind het. Vir enige parasitoloog is ’n parasiet se lewensiklus dus van uiterste belang. Hierdie algemene beginsel is dan ook die raakpunt tussen die lintwurms wat mense besmet en dié wat ’n verskeidenheid werweldiere besmet.
Lintwurms het ’n indirekte lewensiklus (Figuur 2). Dit beteken daar is twee gashere: ’n primêre of finale gasheer waarin die “volwasse” parasiet (m.a.w. ’n lintwurm skoleks wat aan die dundermwand heg met proglottiede wat daaragter ontwikkel) voorkom en ’n tussengasheer waarin ’n ander stadium, bekend as die larfstadium, blaaswurm of metasestood, voorkom. Kom ons gebruik T. solium as voorbeeld, soos weergegee deur Grove (1990). Die mens dien as primêre gasheer, waar die volwasse lintwurm, wat 2 tot 7 meter lank kan wees, in die dundermkanaal voorkom. Net die parasiet se “kop” of skoleks heg vas en proglottiede ontwikkel direk agter die kop. Die oudste, mees “volwasse” proglottiede is dus agter. In effek is elke proglottied ’n eierfabriek met ’n volledige geslagstelsel. Die “ryp” proglottiede wat lewensvatbare eiers bevat, word dan uitgeskei in die feses (verwys na Figuur 1 wat proglottiede in hondefeses toon). ’n Geskikte tussengasheer moet dan weer die infektiewe materiaal (in dié geval eiers) inneem. In T. solium se geval is dit varke. Wanneer die tussengasheer die lewensvatbare eier inneem, ontwikkel die parasiet tot ’n besmetlike stadium (wat in effek beskryf kan word as ’n “ingestulpte lintwurmkoppie”) binne-in die tussengasheer. Die aard, aantal en lokalisering van hierdie stadium verskil wyd tussen die onderskeie lintwurmspesies, selfs binne die enkele genus Taenia (Bush, Fernández, Esch en Seed 2001). In varke (T. solium) en beeste (T. saginata) is dit individuele “masels” (of sistiserki) wat in spiere voorkom, veral in die tong by varke en hart- en kaakspiere by beeste.
Die toestand word sistiserkose genoem en is ook tipies van T. saginata, T. solium, T. crassiceps, T. ovis, T. hydatigena en T. taeniaeformis (Center for Food Security and Public Health 2005).
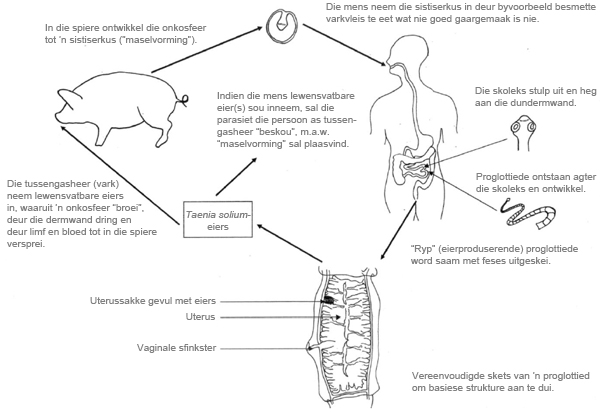
Figuur 2. ’n Tipiese Taenia spp.-lewensiklus, in dié geval Taenia solium.
(Saamgestel deur Dionne Crafford, ClinVet International)
By sommige ander Taenia spp. vorm dit een of meer vloeistofgevulde sakke (senurus) wat honderde koppies kan bevat. Voorbeelde hiervan is T. multiceps en T. serialis (Center for Food Security and Public Health 2005).Soos vroeër vermeld, is dit in die sistserkus-stadium waar veral T. solium-besmetting in mense dodelik kan raak. Wanneer ’n mens die besmetlike eier inneem, “hanteer” die parasiet die persoon as tussengasheer en sal sist-/maselvorming voortgaan. Die eier “broei uit” en die onkosfeer of heksakant-larf dring deur die dermwand en word deur die bloed of limfstelsel na ander liggaamsdele versprei. Die siste kan dus in areas soos die lewer en veral brein vorm, met epilepsie as gevolg (Krecek, Michael, Schantz, Ntanjana, Smith, Dorny, Harrison, Grimm, Praet en Willingham 2008).
In Figuur 3 word ’n soortgelyke situasie uitgebeeld (in dié geval net T. taeniaeformis sistiserki in die lewer van ’n muis).
Die belangrike punt om te onthou is dat ’n mens nie siste sal kry deur lewensvatbare masels in “rou” (m.a.w. ongekookte of halfgaar) varkvleis te eet nie (dit sal ontwikkel tot ’n volwasse wurm in jou dermkanaal). Die mens kan sistiserkose ontwikkel alleenlik deur die inname van lewensvatbare T. solium-eiers (waaruit die onkosfeer sal ontstaan of “uitbroei”) wat dan deur die dermwand sal dring en versprei soos bo beskryf. Vir die lewensiklus om voort te gaan moet ’n geskikte primêre gasheer weer die besmetlike tussenstadium in die tussengasheer inneem.
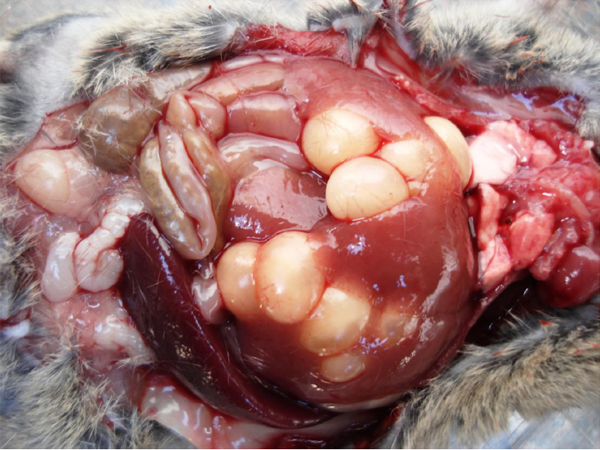
Figuur 3. Taenia taeniaeformis tussenstadiums (sistiserki) binne-in die lewer van ’n muis.
(Foto: Dawie Kok, ClinVet International)
In die geval van T. taeniaeformis sal die kat die besmette muis eet, wat aanleiding sal gee tot volwasse wurms in die dermkanaal (Figuur 4). Om die ander voorbeeld te gebruik wat ons ook reeds genoem het: in T. solium se geval eet mense varkvleis wat lewensvatbare masels bevat (Figuur 2). Die basiese lewensiklus bly dus dieselfde, maar met variasie in die gashere wat betrokke is. In Dipylidium caninum se geval neem honde besmette vlooie in as hulle vlooie probeer verwyder; herkouers en perde neem weer “per ongeluk” grasmyte (wat besmet is met byvoorbeeld Moniezia spp. of Anoplocephala spp. onderskeidelik) in saam met gras. In die dermkanaal stulp die lintwurmkoppies uit, heg aan die dermwand vas en begin proglottiede produseer (sien byvoorbeeld Figuur 4).
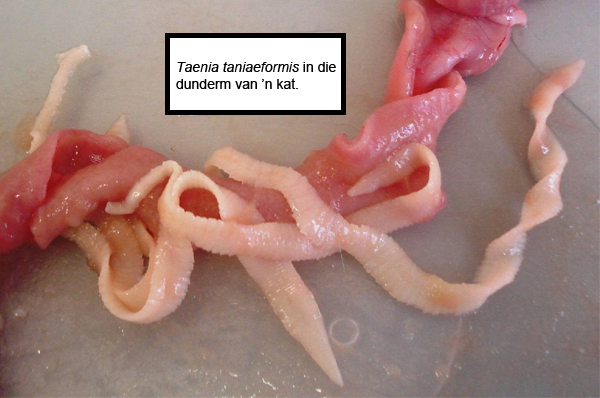
Figuur 4. Taenia taeniaeformis lintwurms uit die dunderm van ’n huiskat.
(Foto verskaf deur Dawie Kok, ClinVet International)
Verster (1969), wat na Taenia spp. wêreldwyd gekyk het, se werk in Suid-Afrika is steeds van onskatbare waarde, soos ook uit ’n baie deeglike oorsig blyk wat deur Loos-Frank (2000) saamgestel is. Van belang in hierdie artikel is slegs spesies wat in Suid-Afrika in honde en katte voorkom.
Uit Verster (1969) se werk blyk dit die volgende spesies te wees: T. hydatigena, T. multiceps, T. ovis ovis, T. pisiformis, T. serialis serialis, T. serialis brauni en T. taeniaeformis. Die laasgenoemde kom in katte (Felidae) voor en die res in hondagtiges (Canidae) (meer inligting oor die betrokke parasiete is in Tabel 2 opgesom).
Al die betrokke verteenwoordigers uit die genus besit twee stelle hake (een stel groot en een stel klein hake) wat gebruik word vir aanhegting aan die dermwand. Die gedeelte van die skoleks wat die hake bevat, staan bekend as die rostellum. Afmetings van hierdie harde strukture (m.a.w. hake) word dan ook grotendeels gebruik om tussen spesies te onderskei.
Die vraag ontstaan of dieselfde Taenia spp.-wurms wat in honde voorkom, ook in “wilde” hondagtiges (soos jakkalse) en katagtiges (soos rooikatte) voorkom in Suid-Afrika, en indien wel, tot watter mate. Die hond (Canis familiaris) het sekerlik nie enige bekendstelling nodig nie. Dit is egter interessant dat “huishoudelike” honde soos ons hulle vandag ken, nie endemies is aan Afrika nie. Argeoloë het die vroegste oorblyfsels van honde in Afrika in Egipte gevind, waar honde na raming sowat 7 000 jaar gelede vir die eerste keer in Afrika uit Europa ingebring is (Gallant 2004). Wat Suid-Afrika betref, dateer die oudste hondoorskot wat al gevind is, tussen 570 en 650 n.C. (Gallant 2004). Die rooijakkals (Canis mesomelas) is aan almal bekend. Dié hondagtige dier word gekenmerk deur die donker “saal” wat oor die rug strek van die nek tot die stert (Walton en Joly 2003). Soos verwag kan word, doen hulle dikwels dieselfde parasietinfeksies (endoparasiete) en infestasies (ektoparasiete) op as honde (Walton en Joly 2003), wat bloedparasietinfeksies insluit (Price en Karstad 1980; Loveridge en Nel 2004). Dit is ook al opgeteken vir ander hondagtiges, soos die rooivos (Vulpes vulpes) (Eira, Vingada, Torres en Miquel 2006) en Arktiese vos (Alopex lagopus) (Skirnisson, Eydal, Gunnarsson en Hersteinsson 1993). Rooijakkalse is geleentheidsvoeders en sal beide prooi vang en op ander beskikbare geskikte oorskot of selfs plantmateriaal aas. Hulle kan dus beskryf word as omnivore, wat ook duidelik blyk uit die struktuur van hulle tande (Skinner en Smithers 1990). Prooi sluit onder andere hase, knaagdiere, insekte en klein herkouers in (Walton en Joly 2003).
Jakkalse, wat sowel in die dag as in die nag aktief kan wees, se gedragspatrone val dan ook dikwels saam met dié van prooidiere (veral knaagdiere) (Skinner en Smithers 1990). Rooikatte (Caracal caracal), daarenteen, is hoofsaaklik jagters en verkies klein tot medium-groot prooi (jong bokke, ander klein soogdiere en voëls) bo aas. Verder is hulle hoofsaaklik naglewend en baie skugter (Skinner en Smithers, 1990).
Om ’n idee te kry van spesies wat algemeen in Suid-Afrika in honde voorkom, is daar na ClinVet International (Edms.) Bpk. se argiefmateriaal gekyk (data wat oorspronklik versamel is van honde in die Grahamstad-omgewing, Oos-Kaap).
Met die hulp van die Vrystaat Sandveld-tak van die Suid-Afrikaanse Jagters- en Wildbewaringsvereniging (SAJWV) is ’n aantal jakkals- en rooikatkarkasse in die Bloemfontein-omgewing (Vrystaat) bekom en vir lintwurms ondersoek. Die geografiese gebiede waaruit monsters geneem is, was dus beperk en is geensins verteenwoordigend van die situasie in Suid-Afrika as geheel nie.
Die hoeveelheid en gehalte van die materiaal was ook nie geskik vir intensiewe statistiese ontledings en breedvoerige uitlatings oor die mate van parasietvoorkoms nie. Nietemin was monsterneming en die versamelde materiaal voldoende om die volgende doelwitte te bereik wat vir die loodsproef gestel is: (1) Identifiseer Taenia spp. wat in honde in Suid-Afrika voorkom en vergelyk dit met Taenia spp. wat in jakkalse voorkom. (2) Identifiseer Taenia spp. wat in rooikatte voorkom en vergelyk dit met wat in honde en jakkalse gekry is. (3) Bespreek variasie in afmetings van aanhegtingshake en bespreek implikasies vir identifikasie van eksemplare wat ondersoek is.
2. Materiaal en metodes
2.1 Versameling van materiaal
Inligting (afmetings van aanhegtingstrukture) oor lintwurms uit honde is verkry uit ClinVet International (Edms.) Bpk. se argiefmateriaal. Lintwurmmateriaal is ook versamel uit jakkalse en rooikatte wat spesifiek vir die studie verwerk is. Materiaal wat gebruik is, is in Tabel 1 hier onder opgesom.
Die dunderm is uit die karkas verwyder en in ’n geskikte houer oopgesny. Sigbare lintwurms is versigtig verwyder met ’n skerppunt-pinset. Die onderliggende, oppervlakkige dermkanaalslymvlies is in sulke gevalle saam met die lintwurmskoleks verwyder om beskadiging van aanhegtingstrukture te beperk. Indien daar talle lintwurms teenwoordig was (of baie jonges, m.a.w. skoleks met min onvolwasse proglottiede aan), is die dermwand liggies geskraap om die mukosa (en die skolekse wat daaraan vasgeheg is) te verwyder. Die skraapsels is dan deur ’n sif met ’n poriegrootte van 150 mikrometer gewas, waarna die parasiete in ’n monsterbottel uitgewas kon word. Parasiete is in 10% formalien gepreserveer.
2.2 Voorbereiding vir meet van aanhegtingshake
Elke wurm (m.a.w. skoleks met proglottiede) is individueel versigtig met ’n skerppunt-pinset uit die monsterbottel verwyder. Die parasiet is vlugtig op handdoekpapier geplaas om dit droog te maak. Daarna is die wurm op ’n mikroskoopglasplaatjie geplaas. Die glasplaatjie is op ’n ligkas geplaas. ’n Skalpel is gebruik om die rostellum wat die hake bevat, van die skoleks af te sny. Die proses is soms ook onder ’n stereomikroskoop uitgevoer vir kleiner wurms. ’n Klein druppel laktofenol (’n ophelderingsvloeistof met ’n suurbasis) is dan op die afgesnyde rostellum geplaas. Die gebruik van suur kan egter die kalkliggaampies aantas en gasborrels veroorsaak. Daar is tydens die ondersoek gevind dat die hake ten spyte hiervan steeds akkuraat gemeet kon word. In toekomstige studies moet Berlese of Hoyer se media egter ook oorweeg word om moontlike probleme in dié verband uit te skakel. Deur gebruik te maak van twee naalde is die rostellum dan onder ’n disseksiemikroskoop gemanipuleer totdat ’n bo-aansig van die aanhegtingshake verkry is. ’n Dekglasie is dan oor die rostellum geplaas. Indien nodig is ’n klein stukkie geskeurde handdoekpapier gebruik om oortollige laktofenol op te droog, waarna die dekglasie se rande met deurskynende naellak geseël is.
2.3 Meet van aanhegtingshake
Die totale lengte van die klein en groot aanhegtingshake onderskeidelik is gebruik vir identifikasiedoeleindes (Figuur 5). Die lengtes van die hake is direk met behulp van ’n mikroskoop-meetoogstuk op ’n 400x-vergroting gemeet. Daar is onderskeidelik vyf groot en vyf klein hake van elke parasiet gemeet, waarna ’n rekenkundige gemiddeld vir elk van die twee stelle bepaal is.
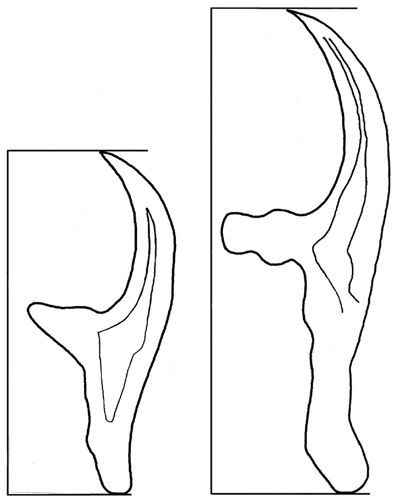
Figuur 5. Grafiese voorstelling van hoe totale lengtes van aanhegtingshake gemeet is.
(Skets: Dionne Crafford, ClinVet International)
3. Resultate
3.1 Materiaal versamel
Die hoeveelheid materiaal wat versamel en ondersoek is, is in Tabel 1 opgesom.
|
Gasheer |
Taenia spp.-wurms |
|||
|
Spesie |
Aantal ondersoek |
Gebied |
Aantal herwin |
Aantal geskik vir neem van afmetings |
|
Hond (Canis familiaris)1 |
8 |
Grahamstad- omgewing |
24 |
24 |
|
Jakkals (Canis mesomelas)2 |
18 |
Bloemfontein- omgewing |
112 |
14 |
|
Rooikat (Caracal caracal)2 |
3 |
Bloemfontein- omgewing |
3 |
2 |
|
1. Data verkry uit ClinVet International (Edms.) Bpk. se argiefmateriaal. 2. Materiaal versamel uit karkasse verkry vanaf die Vrystaat Sandveld-tak van die Suid- |
||||
3.1.1 Taenia spp. waarna verwys word
Besprekings word beperk tot verteenwoordigers van die genus Taenia spp. wat in Suid-Afrika aangeteken is, soos deur Verster (1969) aangedui. In Tabel 2 word meer inligting oor elk van die onderskeie wurmspesies gegee in terme van primêre en tussengashere. Vir grafiese voorstellings is slegs die oorspronklike haaklengtegrense, soos deur Verster (1969) opgeteken, op die onderskeie grafieke aangedui. In Tabel 3 is die afmetings, asook afmetings bygewerk deur Loos-Frank (2000) na ’n volledige literatuuroorsig, egter ook ingesluit om besprekings en vergelykings te vergemaklik.
|
Spesie |
Gashere |
Tussengashere |
Geografiese verspreiding |
|
Taenia hydatigena |
Hondagtiges, Ursus spp. (bere), Felis spp. (Felidae)* |
Meestal herkouers, maar soms ook ander diere, soos varke. |
Wêreldwyd |
|
Taenia multiceps |
Hondagtiges, Hyaena spp. |
Skape, beeste, bokke en ander herkouers, perde en soms mense. |
Wêreldwyd |
|
Taenia ovis ovis |
Hondagtiges |
Skape, bokke en ander herkouers |
Wêreldwyd |
|
Taenia pisiformis |
Hondagtiges, ongereeld katagtiges |
Knaagdiere (veral hase en konyne) |
Wêreldwyd |
|
Taenia serialis serialis |
Hondagtiges, Crocuta spp. |
Meestal hase en konyne (ongereeld ander knaagdiere), karnivore, mense |
Wêreldwyd |
|
Taenia serialis brauni |
Hondagtiges |
Knaagdiere en primate |
Afrika en Amerika |
|
Taenia taeniaeformis |
Katagtiges, |
Knaagdiere, hase/konyne, mense, insekvreters |
Wêreldwyd |
|
* Verster (1969) het egter aangedui dat, gebaseer op ’n literatuuroorsig, huiskatte nie geskikte gashere vir die betrokke spesie is nie. |
|||
|
Spesie |
Afmetings (in mikrometer) |
|||||||
|
Verster (1969) |
Loos-Frank (2000) |
|||||||
|
Klein hake |
Groot hake |
Klein hake |
Groot hake |
|||||
|
Min |
Maks |
Min |
Maks |
Min |
Maks |
Min |
Maks |
|
|
Taenia hydatigena |
118 |
143 |
191 |
218 |
110 |
168 |
169 |
235 |
|
Taenia multiceps |
98 |
136 |
157 |
177 |
73 |
160 |
120 |
190 |
|
Taenia ovis ovis |
111 |
127 |
170 |
191 |
89 |
157 |
131 |
202 |
|
Taenia pisiformis |
128 |
151 |
220 |
261 |
114 |
177 |
200 |
300 |
|
Taenia serialis serialis |
107 |
123 |
154 |
175 |
63 |
129 |
110 |
184 |
|
Taenia serialis brauni |
91 |
102 |
125 |
148 |
47 |
114 |
85 |
160 |
|
Taenia taeniaeformis |
210 |
261 |
370 |
402 |
187 |
293 |
300 |
530 |
|
Let wel: Die minimum- en maksimum-afmetings dui in alle gevalle reikwydtes aan. Hierdie reikwydtes het aansienlik vergroot in die oorsig van Loos-Frank (2000), aangesien mates soos weergegee deur verskeie skrywers in ag geneem is. Hierdie wye reikwydtes vergroot uit die aard van die saak die probleem van oorvleueling in afmetings (en daarmee saam identifikasie van spesies). |
||||||||
3.2 Honde (Canis familiaris) as gasheer
Figuur 6 toon die verspreiding van aanhegtingshaakwaardes in terme van die gemiddelde lengte van klein hake teenoor dié van groot hake vir wurms wat uit honde versamel is. Die reikwydtes deur Verster (1969) aangedui vir die onderskeie Taenia spesies is as blokke aangedui om bespreking te vergemaklik.
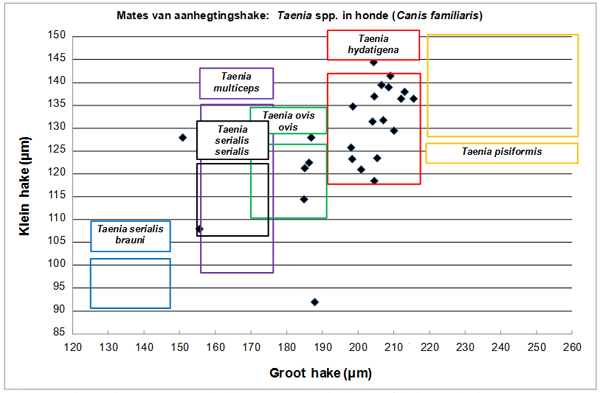
Figuur 6. Grafiese voorstelling van mates van aanhegtingshake van Taenia spp. uit honde soos verkry uit die ClinVet International-argief, saam met reikwydtes aangedui in blokke vir die onderskeie spesies soos weergegee deur Verster (1969).
3.3 Jakkalse (Canis mesomelas) as gasheer
Figuur 7 toon die verspreiding van aanhegtingshaakwaardes in terme van die gemiddelde lengte van klein hake teenoor dié van groot hake vir wurms wat uit jakkalse versamel is. Die reikwydtes deur Verster (1969) aangedui vir die onderskeie Taenia spesies is as blokke aangedui.

Figuur 7. Grafiese voorstelling van mates van aanhegtingshake van Taenia spp. uit jakkalse soos gemeet in die huidige studie, saam met reikwydtes aangedui in blokke vir die onderskeie spesies soos weergegee deur Verster (1969).
Besmettingsstatistieke word hier onder opgesom:
Tabel 4. Besmettingstatistieke vir Taenia spp.-wurms wat uit jakkalse versamel is.
|
Aantal diere ondersoek |
Aantal diere besmet |
Totale aantal Taenia spp.- wurms versamel |
Persentasie besmetting |
Gemiddelde besmettings-intensiteit |
Gemiddelde besmettings-moontlikheid |
|
18 |
4 |
112 |
22,22 |
28,00 |
6,22 |
3.4 Rooikatte (Caracal caracal) as gasheer
Figuur 8 toon die verspreiding van aanhegtingshaakwaardes in terme van die gemiddelde lengte van klein hake teenoor die van groot hake vir wurms wat uit rooikatte versamel is.
Die reikwydtes deur Verster (1969) aangedui vir die onderskeie Taenia spesies is as blokke aangedui om bespreking te vergemaklik.
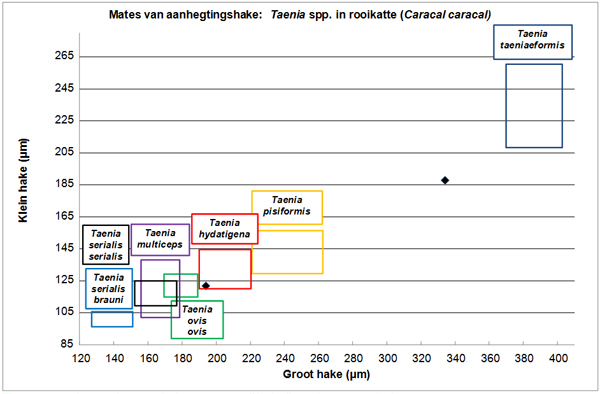
Figuur 8. Grafiese voorstelling van mates van aanhegtingshake van Taenia spp. uit rooikatte soos gemeet in die huidige studie, saam met reikwydtes aangedui in blokke vir die onderskeie spesies soos weergegee deur Verster (1969).
Besmettingstatistieke word hier onder opgesom:
|
Aantal diere ondersoek |
Aantal diere besmet |
Totale aantal Taenia spp.-wurms versamel |
Persentasie besmetting |
Gemiddelde besmettings-intensiteit |
Gemiddelde besmettings-moontlikheid |
|
3 |
2 |
3 |
66,67 |
1,50 |
1,00 |
4. Bespreking
4.1 Materiaal versamel
Soos in Tabel 1 gesien kan word, was ’n groot persentasie van die wurms wat uit jakkalse herwin is, nie bruikbaar nie. Dit was te wyte aan ’n verlies van die aanhegtingstrukture. In 35 uit 115 wurms was alle hake afwesig. In 64 uit 115 wurms was alle groot hake afwesig. Baie van die diere kon eers meer as 12 ure na die diere geskiet is, afgehaal word. In die meeste gevalle was geen verkoelingsfasiliteite beskikbaar nie. Die lintwurmweefsel is baie sag en weefselafbreking begin baie gou. Die gevolg is dat die weefsel van die skoleks en rostellum begin sag raak, sodat die harde hake uittrek en los in die jakkals se dermwand agterbly.
Met ondersoek van die materiaal kon duidelik gesien word waar die hake wel in die rostellum ingebed was (Figuur 9).
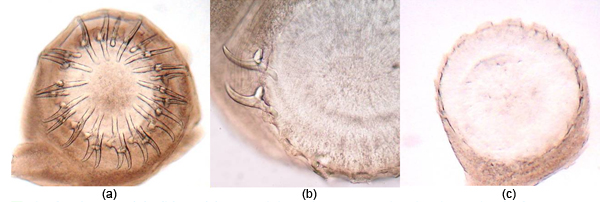
Figuur 9. Voorbeelde van die grade van aanhegtingshaak-verlies in Taenia spp.-lintwurms na herwinning uit jakkalse:
(a) Geen verlies – beide klein en groot aanhegtingshake is teenwoordig (tipies van vars materiaal).
(b) Gedeeltelike verlies – in die meeste gevalle was al die groot hake en die meeste van die klein hake afwesig.
(c) Algehele verlies – al die groot en klein hake is afwesig.
In beide gevalle (b) en (c) is die posisies waar die hake wel ingebed was, duidelik sigbaar. Verlies van hake is opgemerk in jakkalse wat al ’n geruime tyd (in sommige gevalle tussen 12 en 24 uur) dood was voor lintwurmherwinning gedoen is. (Foto’s: Dionne Crafford, ClinVet International)
Alhoewel dit nie gekwantifiseer is nie, was daar ’n opvallende korrelasie tussen die tydperk voor wurmherwinning en die mate van aanhegtingshaak-verlies (Crafford, persoonlike waarneming).
In toekomstige studies sou die ideaal wees om saam met die jagters uit te gaan veld toe, onmiddellik die spysverteringskanaal te verwyder, koel te hou (byvoorbeeld op ys in ’n koelhouer) en so gou moontlik te gaan verwerk in die laboratorium (of ander geskikte area waar water beskikbaar is).
4.2 Honde (Canis familiaris) as gasheer
Sommige vorige studies op Taenia spp.-lintwurmbesmettings in beide tussengashere (bv. njalas Boomker, Horak en Flamand 1991) en primêre gashere (bv. honde – Minnaar, Krecek en Rajput 1999; Minnaar en Krecek 2001; Minnaar, Krecek en Fourie 2002) verwys net na “Taenia sp.”, en identifikasie tot op spesievlak is nie gedoen nie. Uit Figuur 6 is dat duidelik dat die oorgrote meerderheid Taenia-wurms (17 uit 24) wat uit honde versamel is, bes moontlik tot die spesie T. hydatigena behoort.
Die spesie is wêreldwyd baie algemeen en is al gevind in honde in Suid-Afrika (Verster 1979) en Nigerië (Umeche en Hogan 1989); in wolwe (Canis lupis) in Spanje (Segovia, Guerrero, Torres, Miquel en Feliu 2003) en Griekeland (Papadopoulos, Himonas, Papazahariadou en Antoniadou-Sotiriadou 1997); in vosse (Vulpes vulpes) in Griekeland (Papadopoulus e.a. 1997); en in bere (Ursus americanus)in Kanada (Dies 1979), om maar net ’n paar voorbeelde te noem. Die tussenstadiums (sistiserki) kom meestal in herkouers as tussengasheer voor (Tabel 3) en is al herwin uit onder andere skape en bokke in Iran (Radfar, Tajalli en Jalalzadeh 2005); wildsbokke (Odocoileus virginianus) (Schurr, Rabalais en Terwilliger 1988) en skape (Ovis canadensis californiana) (Kistner, Matlock, Wyse en Mason 1977) in Amerika; en boerbokke (Boomker, Horak en Ramsay 1994) en duikers (Sylvicapra grimmia) (Boomker, Keep en Horak 1987) in Suid-Afrika. Die parasiet kan egter ook ander diere as tussengasheer gebruik, soos wilde varke in Spanje (De-la-Muela, Hernández-de-Luján en Ferre 2001) en ape in Soedan (Sulaiman, Williams en Wu 1986).
Dit wil ook voorkom asof minstens vier van die wurms uit honde waarskynlik tot die spesie T. ovis ovis behoort. Beide T. hydatigena en T. ovis kom algemeen in skape (as tussengasheer) voor (Lawson 1994). In veral landelike gebiede is die grootste bron van infektiewe materiaal vir honde bes moontlik afval wat deur mense vir die honde gevoer word. Dit is baie moontlik dat stukke weefsel (bv. spiere vir T. ovis en lewer vir T. hydatigena) wat juis siste bevat, uitgesny sal word en weggegooi sal word in die slagproses (bv. skape en bokke) waar honde dit kan optel en eet. Honde was dus waarskynlik ’n bron van besmetting vir beide T. hydatigena en T. ovis ovis in die landelike gebied met informele behuising in hierdie studie.
Daar is verder een wurm wat moontlik T. serialis serialis of T. multiceps kan wees.
Taenia serialis gebruik meestal hase as tussengasheer (Keith, Cary, Yuill en Keith 1985; Pfaffenberger en Valencia 1988; Whitney 1999), terwyl T. multiceps gewoonlik in skape se breine siste vorm (De Wet en Bath 1994). Aangesien beide die wurms egter blaassiste in die tussengasheer vorm, sou mens baie hoër getalle verwag, aangesien dié tipe siste letterlik honderde besmetlike ingestulpte lintwurmkoppies kan bevat.
Daar is ook twee wurms wat glad nie in Verster (1969) se raamwerk pas nie. Variasie in afmetings en oorvleueling tussen spesies word later bespreek.
4.3 Jakkalse (Canis mesomelas)
Soos aangetoon in Figuur 7, is twee van die wurms wat uit jakkalse versamel is, waarskynlik T. hydatigena. Jakkalse kan maklik die sistiserki inkry deur die ingewande van skape wat pas dood is, of weefsel wat weggegooi is, te vreet. Wat die ander wurms wat moontlik in jakkalse kan voorkom aanbetref, verydel die oorvleueling in haakafmetings tussen T. ovis ovis, T. multiceps en T. serialis serialis enige primêre gevolgtrekkings.
Dit wil voorkom of minstens een parasiet wel T. ovis ovis kan wees, terwyl agt bes moontlik tot T. multiceps of T. serialis serialis kan behoort. Die jakkalse wat ondersoek is, is juis gejag weens skade wat aan skaaptroppe aangerig is, veral tydens lamtyd. Dit is dus baie moontlik dat die wurms wel T. multiceps kan wees, aangesien die parasiet blaassiste in skaapbreinweefsel vorm en inname van so ’n blaas dus tot ’n hele aantal wurms kan lei (hoë gemiddelde intensiteit van besmetting, Tabel 4). Die voorkoms van hierdie parasiet in lammers behoort redelik laag te wees, aangesien die lammers nog nie veel blootstelling aan besmette tussengashere sou hê nie. Daar is egter al getoon dat die druk op die skedelkas weens sistvorming, kan lei tot verdunning van die skedel (Gunn en Pitt 2012), wat sal beteken dat die jakkals wel deur die skedel van volwasse skape sal kan byt. Taenia multiceps is ook al in Australiese wildehonde (dingo’s oftewel Canis latrans) gekry (Henke, Pence en Bryant 2002).
Beide jakkalse en rooikatte voed op klein soogdiere. Hase behoort dus deel uit te maak van die dieet van beide spesies. Taenia pisiformis gebruik hase as tussengasheer (Keith, Keith en Cary 1986; Kietzmann en Hugghins 1986; Pinto, Gomes, Menezes, Gomes en Noronha 2004). Die feit dat geen T. pisiformis in die studie versamel is nie, dui moontlik op ’n lae voorkoms van die parasiet in Suid-Afrika.
4.4 Rooikatte (Caracal caracal)
Uit drie Taenia spp.-lintwurms wat uit twee rooikatte versamel is, was slegs twee geskik om hake van te meet. Figuur 8 toon dat een van die wurms bes moontlik T. hydatigena is.
Uit die huidige literatuurstudie blyk dit dat die voorkoms van die parasiet in rooikatte nog nie in enige publikasies bevestig is nie. Loos-Frank (2000) verwys slegs na die genus Felis in haar gasheerlys, terwyl Verster (1969) meld dat die spesie nie in katte voorkom nie.
Die ander wurm is moontlik T. taeniaeformis, aangesien die hake baie langer is as dié van enige van die Taenia spp.-lintwurms wat in honde voorkom. Die afmetings val wel nie in die reikwydtes wat deur Verster (1969) aangeteken is vir T. taeniaeformis nie, maar wel in die reikwydtes vir die spesie soos aangeteken deur Loos-Frank (2000). Aangesien geen van die ander Taenia-spesies sulke groot hake het nie, kan met redelike sekerheid gesê word dat die wurm wel T. taeniaeformis is. Laasgenoemde is die enigste Taenia-spesie waarvan die volwasse parasiet in huiskatte voorkom. Die stadium van T. taeniaeformis wat infektief is vir katte, kom gewoonlik in die lewer van die tussengasheer (muise en rotte) voor (Theis en Schwab 1992; Seong, Huh, Lee en Oh 1995; Bowman, Barr, Hendrix en Lindsay 2003; Fisher en McGarry 2006). Delahay, Daniels, Macdonald, McGuire en Balharry (1998) het gevind dat die voorkoms van T. taeniaeformis in wilde katte (Felis silvestris) en huiskatte (Felis catus) in Skotland geensins verskil het nie.
Die parasiet kom ook algemeen in huiskatte in Suid-Afrika voor (bv. Baker, Lange, Verster en Van der Plaat 1989). Verder is T. taeniaeformis ook al in Amerikaanse wildekatte (die “bobcat” oftewel Felis rufus) gekry (Heidt, Rucker, Kennedy en Baeyena 1988).
Uit die huidige studie wil dit voorkom of rooikatte in Suid-Afrika Taenia spp.-lintwurms kan huisves wat eie is aan beide honde en katte, ’n voorlopige resultaat wat vir seker in verdere studies ondersoek kan word.
5. Taenia spp.-hake: variasies, neigings en alternatiewe
Die gebruik van harde parasietaanhegtingstrukture se morfometriese afmetings en vorm vir spesie-identifikasie kom algemeen voor. ’n Goeie voorbeeld is dié van ’n groep platwurms (Monogenea) wat op visse voorkom (Pariselle, Bilong Bilong en Euzet 2003; P?ikrylová en Gelnar 2008; Musilová, ?ehulková en Gelnar 2009). Wat Taenia spp. betref, is haaklengte ook een van die primêre veranderlikes wat gebruik word om spesies van mekaar te onderskei. Uit Figure 6 tot 8 staan een ding egter uit soos ’n paal bo water: variasie in afmetings, en daarmee saam oorvleueling tussen spesies. Wanneer die afmetings wat deur Verster (1969) gemaak is, vergelyk word met afmetings wat opgeteken is deur ’n aantal navorsers, soos opgesom deur Loos-Frank (2000) (Tabel 2), word die reikwydtes nog groter. Die vraag ontstaan of variasie gemeet werklik verteenwoordigend is van biologiese/morfologiese verskille, of die gevolg is van variasie in akkuraatheid van mates. Wanneer mates direk met behulp van ’n meetoogstuk gedoen word, sal akkuraatheid afhang van korrekte kalibrasie van die oogstuk.
Wanneer daar fyn verskille tussen strukture is, kan kalibrasie deur verskillende navorsers (en dus ook die mate van akkuraatheid) effens verskil, wat daartoe sal lei dat die mates nie noodwendig direk vergelykbaar is nie. In die huidige studie is van ’n meetoogstuk gebruik gemaak omdat die metode ook deur vorige navorsers gebruik is. In toekomstige studies moet die mates aangevul word deur digitale mates wat direk van die mikroskoopbeeld digitaal op ’n rekenaarskerm gedoen kan word. Laasgenoemde metode is meer akkuraat en sal vergelyking met die histories meer tradisionele metode (en data wat daaruit voortspruit) moontlik maak. Die hake van Taenia sp. is egter redelik groot en stewig (robuust) in vergelyking met aanhegtingstrukture van ander parasietgroepe wat ook gebruik word vir taksonomiese doeleindes. Dit is dus onwaarskynlik dat die verskille in meetmetodes soos bespreek werklik ’n noemenswaardige effek sal hê. Buiten die kalibrasiemetode kan beide navorserondervinding en voorbereiding van materiaal ’n noemenswaardige invloed hê. In dié verband is die dikte van die rostellum en die posisie daarvan op die mikroskoopplaatjie van belang. Indien dit te dik is en/of nie heeltemal plat monteer word nie, kan dit lei tot ’n parallaksfout (slegs hake wat heeltemal horisontaal lê, kan akkuraat gemeet word).
Variasie is ook nie ’n vreemde neiging wanneer dit by die afmetings van aanhegtingstrukture in ander parasietgroepe kom nie. In die genus Gyrodactylus (ook deel van die Monogenea) is daar ook groot variasies in afmetings van harde aanhegtingshake opgeteken (Hanek en Furtado 1973; Harris 1998). Daar is oënskynlik sterk korrelasies tussen hierdie variasie en watertemperatuur (Mo 1991a, 1991b, 1993; Hodnelen en Nilsen 1994; Appleby 1996) en gasheertipe in terme van primêre en sekondêre gashere (Dmitrieva en Dimitrov 2002). Variasie in aanhegtingstruktuurvorm kan ook ’n gevolg wees van die mate waarin die parasiet gasheerspesifiek is (Jarkovský, Moren, Šimková en Gelnar 2004). Weens hierdie variasie in die afmetings a.g.v. geografiese verspreiding of verskille in gasheerspesies word daar nou meer algemeen van molekulêre/genetiese ondersoeke gebruik gemaak.
Dikwels word morfologiese ondersoeke (wat afmetings insluit) gekombineer met molekulêre werk in die genus Gyrodactylus (Šimková, Moren, Jobet, Gelnar en Verneau 2004; Šimková, Mat?jusová en Cunningham 2006; P?ikrylová, Mat?jusová, Jarkovský en Gelnar 2008; P?ikrylová, Mat?jusová, Musilová en Gelnar 2009). Sommige outeurs voel egter tereg dat molekulêre ondersoeke van groot hoeveelhede individuele wurms in ekologiese studies heeltemal onprakties is (Blazek, Bagge en Valtonen 2008). Om dié rede is daar ook ’n aantal addisionele morfometriese afmetings bygevoeg om die sensitiwiteit van onderskeiding te verhoog (sien byvoorbeeld Mizelle en Kritsky 1967).
Uit die huidige loodsstudie is raakpunte tussen die genera Taenia en Gyrodactylus duidelik wat betref variasie in aanhegtingstruktuur-afmetings wat positiewe identifikasie tot op spesievlak bemoeilik.
Om dié rede word daar dikwels ook na ’n aantal ander strukture en veranderlikes gekyk om verder tussen Taenia-spesies te onderskei (sien die baie volledige oorsig deur Loos-Frank 2000 en Hoberg, Jones, Rausch, Eom en Gardner 2000). Daar is ook al heelwat genetiese en molekulêre werk op ’n groot aantal Taenia-spesies gedoen. Hierdie pogings is veral op diagnose van infeksies in mense toegespits (Abraham, Pardini, Vaz, Livramento en Machado 2004; Ogunremi, MacDonald, Geerts en Brandt 2004), m.a.w. T. solium (Rishi en McManus 1988; Chapman, Vallejo, Mossie, Ortiz, Agabian en Flisser 1995; Gasser, Zhu en McManus 1999; von Nickisch-Rosenegk, Silva-Gonzalez en Lucius 1999; Park, Yun, Chung, Kong en Cho 2000; Verastegui, Gilman, Garcia, Gonzalez, Arana, Jeri, Tuero, Gavidia, Levine, Tsang en die sistiserkose-werksgroep in Peru 2003; Yang en Chung 2004) en T. saginata (Chapman e.a. 1995; Von Nickisch-Rosenegk e.a. 1999; Gasser e.a. 1999; Gonzalez, Bonay, Benitez, Ferrer, Harrison, Parkhouse en Garate 2007). Daar is egter ook molekulêre en genetiese werk gedoen (met die oog op spesie-identifikasie) vir ander Taenia-spesies, insluitend T. hydatigena (Allsop, Jones, Allsop, Newton en Macpherson 1987; Gasser e.a. 1999; Von Nickisch-Rosenegk e.a. 1999), T. serialis (Allsop e.a 1987; Gasser e.a. 1999; Von Nickisch-Rosenegk e.a. 1999), T. madoquae (Jones, Allsopp, Macpherson en Allsop 1988; Zhang, Hu, Jones, Allsop, Beveridge, Schindler en Gasser 2007; ), T. regis (Allsop e.a. 1987; Zhang e.a. 2007), T. taeniaeformis (von Nickisch-Rosenegk e.a. 1999; Gasser e.a. 1999), T. pissiformis (von Nickisch-Rosenegk e.a. 1999; Gasser e.a. 1999), T. ovis (Von Nickisch-Rosenegk e.a. 1999; Gasser e.a. 1999), T. multiceps (Gasser e.a. 1999), T. crassiceps (Espíndola, De Gaspari, Nakamura en Vaz 2000), T. martis, T. mustelae en T. parva (Von Nickisch-Rosenegk e.a. 1999). Gevolglik is standaardmetodes al redelik goed beskryf (McManus 1990; Kassai 1999; McManus 2006). Die resultate dui daarop dat hierdie metodes wel van groot waarde is om tussen spesies te onderskei.
Indien verdere intensiewe studies oorweeg word, sal daar beslis ook na ander morfologiese veranderlikes gekyk moet word vir Taenia-spesie-identifikasie in kombinasie met molekulêre/genetiese werk om identifikasies te bevestig. Dit sou byvoorbeeld van waarde wees om proglottiede van die anterior, middel en posterior van die wurm te neem om die morfologie en aantal strukture (bv. uterus, uterussakke, testes en vaginale sfinkter oftewel genitale atrium kwantitatief te beskryf. Dit is dan ook veral die genitale atrium wat van groot waarde blyk te wees, soos ook toegepas deur Verster (1969). So ’n benadering is egter nie moontlik by ’n groep soos Gyrodactylus sp. nie, aangesien “sagte” weefsel/strukture te veel morfologiese plastisiteit toon (Shinn e.a. 1996) en dus van geen taksonomiese waarde is nie. Soos vroeër vermeld, blyk mates van harde strukture, gekombineer met ’n molekulêre aanslag, die mees veelsydige metode te wees.
’n Basiese aanslag wat Taenia spp. betref, sou wees om metodes te bevestig/standaardiseer deur te kyk na maklik identifiseerbare metasestodes en dit van daar uit te brei na volwasse lintwurms. Sulke eksperimentele infeksies kan terselfdertyd met molekulêre metodes gekombineer word om te bevestig tot watter mate taksonomiese gevolgtrekkings ooreenkom. ’n Beginpunt kan die sistiserkus van T. hydatigena wees, omdat dit die grootste blaaswurm in skape en bokke is en in die reël in die omgewing van die lewer aangeheg is, wat nie die geval is met ander sistiserki in skape en bokke nie.
6. Gevolgtrekkings
Vir die ondersoek van Taenia spp. moet materiaal so vars as moontlik wees om verlies van aanhegtingshake voor en tydens parasiet-herwinning te voorkom. In die huidige studie het verwerking van jakkalsdunderms getoon dat daar ’n korrelasie is tussen die tydperk wat die dier dood is voor parasiet-herwinning en die graad van haakverlies.
Die gebruik van verskillende ophelderingsmedia (bv. Hoyer se media teenoor laktofenol) kan ook in volgende studies vergelyk word, aangesien ’n suurbasis potensieel die kalkliggaampies kan aantas en tot vorming van gasborrels kan lei.
Daar is inderdaad oorvleueling tussen die Taenia spp.-samestelling van honde en dié van jakkalse (byvoorbeeld T. hydatigena). Die samestelling van spesies kan beïnvloed word deur faktore soos geografiese verspreiding en dieet. In die loodsstudie was die honde en jakkalse nie afkomstig van dieselfde gebied nie. Verdere studies kan moontlik meer gebiedspesifieke opnames vir beide gasheerspesies insluit. Opnames sal ook beplan moet word om monsters uit meer geografiese gebiede te verkry, om sodoende resultate te lewer wat meer verteenwoordigend is van Suid-Afrika as geheel. So ’n studie kan ook gekombineer word met ’n tipe sosio-ekonomiese studie om te kyk na die omvang en rol van informele slagpraktyke in veral landelike gebiede in die lewensiklus van Taenia spp.-parasiete. In dié opsig sal dit veral interessant wees om na gebiede te kyk waar T. multiceps-senuri algemeen in skaapbreine voorkom. Tydens so ’n studie sal ’n poging aangewend moet word om die graad van predasie deur jakkalse te kwantifiseer, die rol van honde in die lewensiklus te evalueer en dit te vergelyk met die voorkoms van die Taenia spp. in beide gashere.
Verder wil dit voorkom of rooikatte lintwurms kan huisves wat eie is aan honde (T. hydatigena), asook wat eie is aan katte (T. taeniaeformis). Die verskynsel moet in verdere studies bevestig en verder bestudeer word.
Verdere morfometriese (ondersoek van hake, maar ook ander strukture se mates deur gebruik te maak van beide oogstuk- en digitale meetmetodes), eksperimentele (deur metodes te ontwikkel/standaardiseer om larfstadiums wat moontlik makliker identifiseerbaar is) en molekulêre (waarvoor metodes reeds bestaan) studies is nodig om dié loodsstudie se tentatiewe gevolgtrekkings verder te ondersoek.
Bibliografie
Abraham, R., Pardini, A. X., Vaz, A.J., Livramento, J.A. en Dos Ramos Machado, L. (2004). Taenia antigens detection in the cerebrospinal fluid of patients with neurocticercosis and its relationship with clinical activity of the disease. Arquivos de Neuro-Psiquiatria: 62(3–B), 756–760.
Allsop, B.A., Jones, A., Allsop, M.T.E.P., Newton, S.D. en Macpherson, C.N.L. (1987). Interspecific characterization of several taeniid cestodes by isoenzyme analysis using iselectric focusing in agarose. Parasitology: 95, 593–601.
Appleby, C. (1996). Variability of the opisthaptoral hard parts of Gyrodactylus callariatis Malmberg, 1957 (Monogenea: Gyrodactylidae) from Atlantic cod Gadus morhua L. in the Oslo Fjord, Norway. Systematic Parasitology: 33, 199–207.
Baker, M.K., Lange, L., Verster, A. en Van der Plaat, S. (1989). A survey of helminthes in domestic cats in the Pretoria area of Transvaal, Republic of South Africa. Part 1: The prevalence and comparison of burdens of helminths in adult and juvenile cats. Journal of the South African Veterinary Association: 60(3), 139–142.
Blažek, R.D., Bagge, A. en Valtonen, E.T. (2008). Monogenean assemblages and the apparent transmission capability of monogeneans between related fish species: an experimental study. Parasitology Research: 102, 1359–1366.
Boomker, J., Horak, I.G. en Flamand, J.R.B. (1991). Parasites of South African wildlife. XII. Helminths of Nyala, Tragelaphus Angasii, in Natal. Onderstepoort Journal of Veterinary Research: 58, 275–280.
Boomker, J. Horak, I.G. en Ramsay, K.A. (1994). Helminth and arthropod parasites of indigenous goats in the northern Transvaal. Onderstepoort Journal of Veterinary Research: 61, 13–20.
Boomker, J. Keep, M.E. en Horak, I.G. (1987). Parasites of South African Wildlife. I. Helminths of Bushbuck, Tragelaphus scriptus, and grey Duiker, Sylvicapra grimmia from the Weza State Forest, Natal. Onderstepoort J. Vet. Res.: 54, 131–134.
Bowman, D.D., Barr, S.C., Hendrix, C.M. en Lindsay, D.S. (2003). Gastro-intestinal parasites of cats. In: Companion and exotic animal physiology. Red.: D.D. Bowman. International Veterinary Information Service, Ithaca NY.
Bush, A.O., Fernández, J.C., Esch, G.W. en Seed, J.R. (2001). Parasitism – The diversity and ecology of animal parasites. Cambridge University Press.
Center for Food Security en Public Health. (2005). Taenia infections. College of Veterinary Medicine, Iowa State University, http://www.cfsph.iastate.edu.
Chapman, A., Vallejo, V., Mossie, K.G., Ortiz, D., Agabian, N. en Flisser, A. (1995). Isolation and characterization of species-specific DNA probes from Teania solium and Tania saginata and their use in egg detection assay. Journal of Clinical Microbiology: 33(5), 1283–1288.
Delahay, R.J., Daniels, M.J., Macdonald, D.W., McGuire, K. en Balharry, D. (1998). Do patterns of helminth paratism differ between groups of wild-living cats in Scotland? Journal of Zoology (London): 245, 175–183.
De-la-Muela, N., Hernández-de-Luján, S. en Ferre, I. (2001). Helminths of wild boar in Spain. Journal of Wildlife Diseases: 37(4),840–843.
De Wet, J. en Bath, G. (1994). Kleinveesiektes. Tafelberg Uitgewers Beperk.
Dies, K.H. (1979). Helminths recovered from Black bears in the Peace River region of Northwestern Alberta. Journal of Wildlife Diseases: 15,49–50.
Dmitrieva, E. en Dimitrov, G. (2002). Variability in the taxonomic characters of Black Sea gyrodactylids (Monogenea). Systematic Parasitology: 51, 199–206.
Eira, C., Vingada, J., Torres, J. en Miquel, J. (2006). The helminth community of the red fox, Vulpes vulpes, in Dunas de Mira (Portugal) and its effects on host condition. Wildlife Biology in Practice: 2(1), 26–36.
Eom, K.S. en Rim, H-J. (1992). Experimental human infection with Asian Teania saginata metacestodes obtained from naturally infected Korean domestic pigs. The Korean Journal of Parasitology: 30(1), 21–24.
Eom, K.S., Rim, H-J. en Geerts, S. (1992). Experimental infection of pigs and cattle with eggs of Asian Teania saginata with special reference to its extrahepatic viscerotropism. The Korean Journal of Parasitology: 30(4), 269–275.
Espíndola, N.M., De Gaspari, E.N., Nakamura, P.M. en Vaz, A.J. (2000). Production of Monoclonal antibodies anti-Taenia crassiceps cysticerci with cross-reactivity with Taenia solium antigens. Revista do Instituto Medicina Tropical de São Paulo: 42(3), 175–177.
Fisher, M. en McGarry, J. (2006). Focus on small animal parasitology. Bayer HealthCare AG, Animal Health Division.
Flisser, A., Viniegra, A-E., Aguilar–Vega, L., Garza-Rodriguez, A., Maravilla, P. en Avila, G. (2004). Portrait of human tapeworms. Journal of Parasitology: 90(4), 914–916.
Gallant, J. (2004). The dog for Africa. Animal talk, September 2004: 18–22.
Galán-Puchades, M.T. en Fuentes, M.V. (2000). The Asian Taenia and the possibility of cysticercosis. The Korean Journal of Parasitology: 38(1), 1–7.
Gasser, R.B., Zhu, X, en McManus, D.P. (1999). NADH dehydrogenase subunit 1 and cytochrome c oxidase subunit I sequences compared for members of the genus Taenia (Cestoda). International Journal for Parasitology: 29, 1965–1970.
Gonzalez, L.M., Bonay, P., Benitez, L., Ferrer, E., Harrison, L.J.S., Parkhouse R.M.E. en Garate, T. (2007). Molecular and functional characterization of a Taenia adhesion gene family (TAF) encoding potential protective antigens of Taenia saginata oncospheres. Parasitology Research: 100, 519–528.
Grove, D.I. (1990). A History of Human Helminthology. Bookcraft (Bath) Ltd.
Gunn, A. en Pitt, S.J. (2012). Parasitology: An integrated approach. Wiley-Blackwell, Suusex, United Kingdom.
Hanek, G. en Furtado, J.I. (1973). Gyrodactylus fernenoi sp. n. (Monogenea: Gyrodactylinae) from Clarias batrachus L. in Malaysia. Zoologischer Anzeiger Leipzig: 191, 3/4, 196–198.
Harris, P.D. (1998). Extreme morphological variation between related individuals of Gyrodactylus pungitti Malmberg, (Monogenea). Systematic Parasitology:39, 137–140.
Harrison, L.J.S., Grimm, F., Praet, N. en Willingham, A.L. (2008). Prevalence of Taenia solium cycticercosis in swine from a community-based study in two villages of the Eastern Cape Province, South Africa. Veterinary Parasitology: 154, 38–47.
Heidt, G.A., Rucker, R.A., Kennedy, M.L. en Baeyena, M.E. (1988). Hematology, intestinal parasites, and selected disease antibodies from a population of Bobcats (Felis rufus) in Central Arkansas. Journal of Wildlife Diseases: 24(1), 180–183.
Henke, S.E., Pence, D.B. en Bryant, F.C. (2002). Effect of short-term coyote removal on populations of coyote helminths. Journal of Wildlife Diseases: 38(1), 54–67.
Hoberg, E.P., Alkire, N.L., De Queiroz, A. en Jones, A. (2001). Out of Africa: origins of the Teania tapeworms in humans. Proceedings of the Royal Society of London, series. B: 268, 781–787.
Hoberg, E.P., Jones, A., Rausch, R.L., Eom K.S. en Gardner, S.L. (2000). A phylogenetic hypothesis for species of the genus Taenia (Eucestoda: Teaniidae). Journal of Parasitology: 86(1), 89–98.
Hodnelen, K. en Nilsen, F. (1994). On the occurrence and morphology of Gyrodactylus pterygialis from Saithe Pollachius virens in a Norwegian Fjord. Journal of Parasitology: 80(6), 938–945.
Jarkovský, J., Moren, S., Šimková, A. en Gelnar, M. (2004). Reproductive barriers between conganderic monogenean parasites (Dactylogyrus: Monogenea): Attachment apparatus morphology or copulatory organ incompatibility? Parasitology Research: 92, 95–105.
Jones, A., Allsopp, B.A., Macpherson, C.N.L. en Allsop, M.T.E.P. (1988). The identity, life cycle and isoenzyme characteristics of Taenia madoquae (Pellegrini, 1950) n. comb. from silver-backed jackal (Canis mesomelas Schreber, 1775) in East Africa. Systematic Parasitology: 11, 31–38.
Kassai, T. (1999). Veterinary Helminthology, Butterworth-Heinemann, Oxford.
Keith, I.M., Keith L.B. en Cary, J.R. (1986). Parasitism in a declining population of snowshoe hares. Journal of Wildlife Diseases: 22(3), 349–363.
Keith, L.B., Cary, J.R., Yuill, T.M. en Keith, I.M. (1985). Prevalence of helminths in a cyclic snowshoe hare population. Journal of Wildlife Diseases: 21(3), 233–253.
Kietzmann, G.E. en Hugghins, E.J. (1986). Helminths of lagomorphs in South Dakota. Journal of Wildlife Diseases: 22(2), 276–278.
Kistner, T.P., Matlock, S.M., Wyse, D. en Mason, G.E. (1977). Helminth parasites of bighorn sheep in Oregon. Journal of Wildlife Diseases: 13, 125–130.
Krecek, R.C., Michael, L.M., Schantz, P.M., Ntanjana, L., Smith, M.F., Dorny, P.
Lawson, J.R. (1994). Hydatid disease and sheep measles: the history of their control and the economics of a recent change of control policy. New Zealand Journal of Zoology: 21, 83–89.
Loos-Frank, B. (2000). An up-date of Verster’s (1969) “Taxonomic revision of the genus Teania Linnacus” (Cestoda) in table format. Systematic Parasitology: 45, 155–183.
Loveridge, A.J. en Nel, J.A.J. (2004). Black-backed jackal, Canis mesomelas Schreber, 1775. In: Canids: Foxes, Wolves, Jackals and Dogs. Reds.: C. Sillero-Zubiri, M. Hoffmann en D.W. Macdonald. IUCN/SSC Canid Specialist Group, The World Conservation Union.
McManus, D.P. (1990). Characterisation of taeniid cestodes by DNA analysis. Revue Scientific et Technique (International Office of Epizootics): 9(2), 489–510.
McManus, D.P. (2006). Molecular discrimination of teaniid cestodes. Parasitology International: 55, S31–S37.
Minnaar, W.N. en Krecek, R.C. (2001). Helmiths in dogs belonging to people in a resource-limited urban community in Gauteng, South Africa. Onderstepoort Journal of Veterinary Research: 68, 111–117.
Minnaar, W.N., Krecek, R.C. en Rajput, J.I. (1999). Helminth parasites of dogs from two resource-limited communities in South Africa. Tydskrif van die Suid-Afrikaanse Veterinêre Vereniging: 70(2), 92–94.
Minnaar, W.N., Krecek, R.C. en Fourie, L.J. (2002). Helminths in dogs from a peri-urban resource-limited community in Free State Province, South Africa. Veterinary Parasitology: 107, 343–349.
Minozzo, J.C. Gusso, R.L.F., De Castro, E.A., Lago, O. en Soccol, V.T. (2002). Experimental bovine infection with Taenia saginata eggs: recovery rates and cysticerci location. Brazilian Archives of Biology and Technology: 45(4), 451–455.
Mizelle, J.D. en Kritsky, D.C. (1967). Studies on monogenetic trematodes. XXXIII. New Species of Gyrodactylus and a key to the North American Species. Transactions of the American Microscopical Society: 86(4), 390–401.
Mo, T.A. (1991a). Variations of opisthaptoral hard parts of Gyrodactylus salaris Malmberg, 1957 (Monogenea: Gyrodactylidae) on rainbow trout Oncorhynchus mykiss (Walbaum, 1792) in a fish farm, with comments on the spreading of the parasite in south-eastern Norway. Systematic Parasitology:20, 1–9.
—. (1991b). Variations of opisthaptoral hard parts of Gyrodactylus salaris Malmberg, 1957 (Monogenea: Gyrodactylidae) on parr of Atlantic Salmon Salmo salar L. in laboratory experiments. Systematic Parasitology: 20, 11–19.
—. (1993). Seasonal variations of the opisthaptoral hard parts of Gyrodactylus derjavini Mikailov, 1975 (Monogenea: Gyrodactylidae) on brown trout Salmo trutta L. parr and Atlantic salmon S. salar L. parr in the River Senvikselva, Norway. Systematic Parasitology:26, 225–231.
Musilová, N., ?ehulková, E. en Gelnar, M. (2009). Dactylogyrids (Platyhelminthes: Monogenea) from the gills of the African carp, Labeo coubie Rüppell (Cyprinidae), from Senegal, with descriptions of three new species of Dactylogyrus and redescription of Dactylogyrus cyclocirrus Paperna, 1973. Zootaxa: 2241, 47–68.
Ogunremi, O., MacDonald, G., Geerts, S. en Brandt, J. (2004). Diagnosis of Taenia saginata cysticercosis by immunohistochemical test on formation-fixed and paraffin-embedded bovine lesions. Journal of Veterinary Diagnostic Investigation: 16, 438–441.
Papadopoulos, H., Himonas, C., Papazahariadou, M. en Antoniadou-Sotiriadou, K. (1997). Helminthes of foxes and other wild carnivores from rural areas in Greece. Journal of Helminthology: 71, 227–231.
Pariselle, A., Bilong Bilong, C.F. en Euzet, L. (2003). Four new species of Cichlidogyrus
Paperna, 1960 (Monogenea: Ancyrocephalidae), all gill parasites from African mouthbreeder tilapias of the genera Sarotherodon and Oreochromis (Pisces, Cichlidae), with a redescription of C. thurstonae Ergens, 1981. Systematic Parasitology: 56, 201–210.
Park, S-K., Yun, D-H., Chung, J-Y., Kong, Y. en Cho, S-Y. (2000). The 10 kDa protein of Taenia solium metacestodes shows genus specific antigenicity. The Korean Journal of Parasitology: 38(3), 191–194.
Pfaffenberger, G.S. en Valencia, V.B. (1988). Helminths of sympatric black-tailed Jack Rabbits (Lepus californicus) and Desert Cottontails (Sylvilagus audubonii) from the high plains of Eastern New Mexico. Journal of Wildlife Diseases: 24(2),375–377.
Pinto, R.M., Gomes, D.C. Menezes, R.C. Gomes, C.T. en Noronha, D. (2004). Helminths of rabbits (Lagomorpha, Leporidae) deposited in the helminthological collection of the Oswaldo Cruz Institute. Revista Brasileira de Zoologia: 21(3), 599–604.
Price, J.E. en Karstad, L.H. (1980). Free-living jackals (Canis mesomelas) – potential reservoir hosts for Ehrlichia canis in Kenya. Journal of Wildlife Diseases: 16(4), 469–473.
P?ikrylová, I. en Gelnar, M. (2008). The first record of Macrogyrodactylus species (Monogenea, Gyrodactylidae) on freshwater fishes in Senegal with the description of Macrogyrodactylus simentiensis sp. nov., a parasite of Polypterus senegalus Cuvier. Acta Parasitologica: 53, 1–8.
P?ikrylová, I., Mat?jusová, I., Jarkovský, J., en Gelnar, M. (2008). Morphometric comparison of three members of the Gyrodactylus nemachili-like species group (Monogenea: Gyrodactylidae) on Barbatula barbatula L. in the Czech Republic, with a reinstatement of G. papernai Ergens & Bychowsky, 1967. Systematic Parasitology: 69, 33–44.
Přikrylová, I., Mat?jusová, I., Musilová, N., Gelnar, M. (2009). Gyrodactylus species (Monogenea: Gyodactylidae) on the cichlid fishes of Senegal, with the description of Gyrodactylus ergensi n. sp. from Mango tilapia, Sarotherodon galilaeus L. (Teleostei: Cichilidae). Parasitology Research: 106(1), 1–6.
Radfar, M.H., Tajalli, S. en Jalalzadeh, M. (2005). Prevalence and morphological characterization of Cysticercus tenuicollis (Teania hydatigena cysticerci) from sheep and goats in Iran. Veterinarski Arhiv: 75(6), 469–476.
Rishi, A.K. en McManus, D.P. (1988). Molecular cloning of Taenia solium genomic DNA and characterization of taeniid cestodes by DNA analysis. Parasitology: 97, 161–176.
Schurr, K., Rabalais, F. en Terwilliger, W. (1988). Cysticerus tenuicollis: A new state record for Ohio. Ohio Journal of Science: 88(3), 104–105.
Segovia, J-M., Guerrero, R., Torres, J., Miquel, J. en Feliu, C. (2003). Ecological analyses of the intestinal helminth communities of the wolf, Canis lupus, in Spain. Folia Parasitologica: 50, 231–236.
Seong, J.K., Huh, S., Lee, J-S. en Oh, Y-S. (1995). Helminths in Rattus norvegicus captured in Chunchon, Korea. The Korean Journal of Parasitology: 33(3), 235–237.
Shinn, A.P., De Clers, S., Gibson, D.I. and Sommerville, C. (1996). Multivariate analyses of morphometrical features from Gyrodactylus spp. (Monogenea) parasitising British salmonids: Light microscope based studies. Systematic Parasitology: 33, 115–125.
Šimková, A., Mat?jusová, I. en Cunningham, C.O. (2006). A molecular phylogeny of the Dactylogyridae sensu Kritsky & Boeger (1989) (Monogenea) based on the D1–D3 domains of large subunit rDNA. Parasitology: 133, 43–53.
Šimková, A., Moren, S., Jobet, E., Gelnar, M. en Verneau, O. (2004). Molecular Phylogeny of conganderic monogenean parasites (Dactylogyrus): A case of intrahost speciation. Evolution: 58(5), 1001–1018.
Skinner, J.D. en Smithers, R.H.N. 1990. The Mammals of the Southern African Subregion, Tweede uitgawe, Universiteit van Pretoria, Pretoria.
Skirnisson, K. Eydal, M. Gunnarsson, E. en Hersteinsson, P. (1993). Parasites of the arctic fox (alopex lagopus) in Iceland. Journal of Wildlife Diseases: 29(3), 440–446.
Sulaiman, S., Williams, J.F. en Wu, D. (1986). Natural infections of Vervet Monkeys (Cercopithecus aethiops) and African Red Monkeys (Erythrocebus patas) in Sudan with taeniid cysticerci. Journal of Wildlife Diseases: 22(4), 586–587.
Theis, J.H. en Schwab, R.G. (1992). Seasonal prevalence of Taenia taeniaeformis: relationship to age, sex, reproduction and abundance of an intermediate host (Persmyscus maniculatus). Journal of Wildlife Diseases: 28(1), 42–50.
Umeche, N. en Hogan, U.E.E. (1989). A study of intestinal helminthes of dogs in Calabar. Nigeria. Archivio Veterinario Italiano: 40(2), 128–131.
Verastegui, M., Gilman, R.H., Garcia, H.H., Gonzalez, A.E., Arana, Y., Jeri, C., Tuero, I., Gavidia, C. M., Levine, M., Tsang, V.C.W. en The Cysticercosis Working Group in Peru. (2003). Prevalence of antibodies to unique Taenia solium oncosphere antigens in taeniasis and human and porcine cysticercosis. American Journal of Tropical Medicine and Hygiene: 69(4), 438–444.
Verster, A. (1969). A taxonomic revision of the genus Teania Linnaeus, 1758. S Str. Onderstepoort Journal of Veterinary Research: 36(1), 3–58.
—. (1979). Gastro-intestinal helminths of domestic dogs in the Republic of South Africa. Onderstepoort Journal of Veterinary Research: 46, 79–82.
Walton, L.R. en Joly, D.O. (2003). Canis mesomelas. Mammalian Species:715:1–9 (30 Julie 2003 deur die American Society of Zoologists gepubliseer).
Wang, I.C., Chung, W.C., Lu, S.C. en Fan, P.C. (2000). Rodent model for long-term maintenance and development of the viable cysticerci of Teania saginata asiatica. The Korean Journal of Parasitology: 38(4), 237–244.
Whitney, H. (1999). Common diseases of the snowshoe hare (1): Tapeworm cysts. Publication AP007, Department of Forest Resources and Agrifoods, St. John’s, NF.
Yang, H-J. en Chung, Y-B. (2004). Immunolocalization of the 150 kDa protein in cyst fluid of Taenia solium metacestodes. The Korean Journal of Parasitology: 42(2), 81–84.
Zhang, L., Hu, M., Jones, A., Allsopp, B.A., Beveridge, I., Schindler, A.R. en Gasser, R.B. (2007). Characterization of Teania madoquae and Taenia regis from carnivores in Kenya using genetic markers in nuclear and mitochondrial DNA, and their relationship with other selected teaniids. Molecular and Cellular Probes: 21, 379–385.
Bedankings
Vrystaat Sandveld-tak van die Suid-Afrikaanse Jagters- en Wildbewaringsvereniging (SAJWV) (in besonder Danie Lombaard) vir materiaal en logistieke hulp.
Die eerste skrywer se vrou, Nicolette, wat gedwee en geduldig jakkalskarkasse in haar kar se kattebak moes aanry ...
ClinVet International (Edms.) Bpk. vir gebruik van fasiliteite en logistieke hulp.